![]()
![]()
![]()
![]()
**To view all of the graphics in this site you must use Internet Explorer. 5.0 +**
FoF1-ATP Synthase
The research in our lab primarily concerns the FoF1-ATP synthase that is responsible for ATP synthesis in all known living organisms.
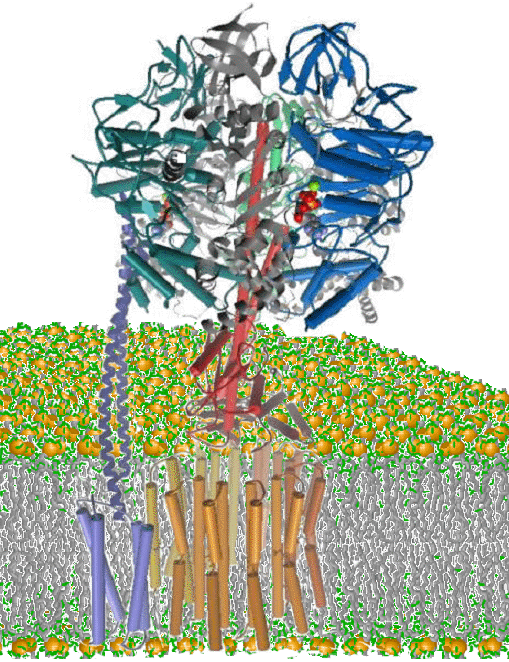
The following is meant to serve as a primer for anyone who would like to learn the basic function of the FoF1-ATP synthase in photosynthetic organisms. To see an example of ongoing projects in our lab visit the research projects page by following the link on the left.
INTRODUCTION
The energy to drive the metabolic processes of all life forms on earth is ultimately derived from photosynthetic processes. The principle means used by a living organism to store this energy is through the production of adenosine 5’-triphosphate (ATP) or the reduced form of nicotinamide adenine dinucleotide (NADH) and its phosphorylated variant nicotinamide adenine dinucleotide phosphate (NADPH) that is used in photosynthetic processes. Due to the wide variety of metabolic processes that use ATP as an energy source, this molecule is often referred to as the energy currency of a cell. Energy stored in the high-energy phosphoryl bond of ATP is released when this bond is cleaved by an ATPase reaction to form ADP and inorganic phosphate (Pi). The energy stored in reduced NADPH derives from its negative redox potential relative to other cellular components.
During photosynthesis, ATP is synthesized from ADP and Pi by the chloroplast FoF1 ATP synthase. This enzyme is composed of the thylakoid membrane-embedded Fo protein complex, and the attached F1 protein complex that protrudes into the aqueous stroma of the chloroplast (Figure 1). The Fo is composed of subunits a, b, b’, and c (also known as subunits IV, I, II, and III, respectively) while F1 contains subunits a, b, g, d, and e.
Involvement of Photosynthetic Light
Reactions.
The energy used by FoF1 to drive the synthesis of ATP from ADP and Pi is derived from the capture of light by light-harvesting and reaction center protein complexes in thylakoid membranes (1). Once absorbed, light energy is transferred to Photosystem I (PSI) or Photosystem II (PSII) where it induces oxidation of the reaction center chlorophyll and reduction of an electron acceptor that is far more electronegative than the ground state chlorophyll. Since molecules with a more electronegative redox potential store more energy upon reduction, the reduction of the initial electron acceptors in PSI and PSII can be considered to be the step in which light energy is first captured as chemical energy during photosynthesis. Due to the instability of this initial product, a series of sequential redox reactions occurs to prevent the recombination of charges in the reaction center chlorophyll and the initial electron acceptor. These reactions result in the reduction of a molecule that is less electronegative than the initial acceptor, but still substantially more electronegative than the reaction center chlorophyll. The oxidized PSII reaction center abstracts electrons from water to release molecular oxygen and protons into the thylakoid lumen.
Reduction of plastoquinone by PSII consumes two protons from the stroma. The reduced, lipid soluble quinol is then oxidized by the cytochrome b6/f protein complex in the thylakoid which results in the deposition of the protons in the thylakoid lumen. The reducing equivalents then transfer to oxidized PSI reaction center chlorophyll. The reducing equivalents generated by PSI reduce the iron-sulfur cluster of ferredoxin, a water-soluble protein in the stroma. Ferredoxin in turn reduces NADP+ via the enzyme ferredoxin NADP+ reductase (FNR), a peripheral membrane protein on the stromal side of the thylakoid that is associated with an intrinsic protein in the thylakoid. The net result of the photosynthetic electron transfer reactions is that electrons from water with a redox potential of +0.81V are used to reduce NADP+ that has a redox potential of –0.32V that accompanies vectorial movement of protons from the stroma to the thylakoid lumen (1).
Chemiosmotic
Energy Coupling
Some of the energy initially captured by the photosynthetic light reactions is used to power the vectorial movement of protons from the stroma to the thylakoid lumen, thereby forming a transmembrane proton gradient (3). This proton translocation causes a small pH increase of the stroma, and a much larger pH decrease in the lumen due to its relatively small volume. This leads to the non-equilibrium condition of a pH difference of 3 to 4 units across the thylakoid membrane during the photosynthetic light reactions. The rate that protons escape from the lumen to the stroma is relatively fast (t1/2 of decay of the trans-membrane proton gradient is ~1 sec). Consequently, the instability of the proton gradient makes it unsuitable for long-term energy storage. Instead, the proton gradient is used by the FoF1 ATP synthase as the energy source to drive ATP synthesis. In this manner, the photosynthetic light reactions are coupled to the synthesis of ATP via the proton gradient. Reagents like ammonia that can transport protons across the membrane more rapidly than the FoF1 ATP synthase will inhibit ATP synthesis because they collapse the proton gradient. These reagents that uncouple ATP synthesis from electron transfer reactions, called uncouplers, typically increase the rate of the latter reactions by relieving back-pressure from the proton gradient.
Energy coupling occurs because Fo serves as an efficient conduit to move protons across the thylakoid membrane and back toward equilibrium with the stroma. Protons move to the stroma in response to the energy gradient that is derived from the concentration difference across the membrane (DpH). Because each proton also carries a positive charge, the charge difference across the membrane (DY) also contributes to this proton energy gradient. A proton concentration difference of about 1000 fold across the membrane (DpH = 3) provides sufficient energy for FoF1 to drive ATP synthesis. Even though FoF1 transports only protons across the membrane, nonequilibrium concentration gradients of other ions like K+ can contribute to the energy of the proton gradient by changing the DY if the ion is permeable to the membrane.
In this chemiosmotic coupling process the magnitude of the energy gradient, designated the proton-motive force (pmf or mH+), is related to the trans-membrane concentration and charge differences in mV at 30°C by equation 1. The protonmotive force translates into more conventional energy terms because it is the sum of the free energy derived from the trans-membrane concentration difference (equation 2), and the electrical potential gradient generated by the trans-membrane concentration gradient of charged species (equation 3) where n is the charge on the ion (+1 for protons). This relationship simplifies to equation 4.
DmH+
= DY
–
59DpH
(Eq. 1).
DG = 2.3RTlog [H+lumen]/[ H+stroma] = 2.3RTDpH (Eq. 2),
DG = –n
F DE =
–n F
DY
(Eq. 3),
DG = – FDmH+ (Eq. 4).
Alternating Site Mechanism of the FoF1
Biomolecular Motor.
The FoF1 ATP synthase operates as a molecular rotary motor (4). The protein subunits that comprise the rotor and stator of this motor are g, e, c and a, b, d, a, b, b’, respectively. The c subunits assemble into a ring that is currently estimated to contain 14 subunits. Each transmembrane helical c subunit has an aspartic acid that is protonated by the protons of the lumen via a channel located on subunit a. An arginine at a separate location on subunit a displaces the proton from subunit c to release it to the stroma. Each successive proton displacement induces the stepwise rotation of the c subunit ring, driven by the transmembrane proton gradient.
The g and e subunits, docked to the c subunit ring, also rotate in response to the proton gradient-driven step-wise rotation and comprise the rotor of the biomolecular motor. The N and C termini of the g subunit form a coiled-coil that protrudes through a ring composed of three a and b subunit heterodimers that comprise each catalytic site. The ring of ab heterodimers along with subunits a, b, b’, and d comprise the stator of the motor.
Each catalytic site adopts a conformation in response to the rotational position of the g subunit such that the three catalytic sites are in different conformations with one site empty. Completion of a catalytic cycle at any one site requires 360° rotation of the g subunit. In what is known as the binding-change or alternating site mechanism, the binding of a Mg2+-ADP complex and Pi to the empty site triggers a 120° step rotation of the g subunit, driven by the proton gradient, that releases ATP from a different catalytic site (5). The energy from the flux of 3 protons through the membrane is minimally required to drive a 120° rotation and ATP production, although the measured ratio of protons/ATP is ~ 4.
The FoF1 ATP synthase uses the nonequilibrium proton gradient to drive the reaction, ADP + Pi = ATP + H2O, far beyond the point of equilibrium, in favor of the products. Most enzymes that use ATP hydrolysis as a source of energy, derive the energy from the reaction by returning the concentration ratio of ATP/ADP+Pi toward equilibrium. Sequential changes in catalytic site conformation maintain this ratio away from equilibrium. In the initial conformation, the binding of ADP and Pi is preferred to ATP. The first conformational change increases the affinity of the catalytic site for substrates and products, and has a low activation energy barrier for ATP synthesis that allows rapid interconversion of substrates and products. The second conformational change converts the site to one that favors ATP over ADP and Pi, while the third decreases the affinity for ATP so that, even when the ATP/ADP+Pi ratio is high, ATP hydrolysis is minimized and ATP dissociates.
Regulation of ATP Synthesis in
Chloroplasts.
After chloroplasts have reached steady state in the light, the onset of darkness leads to a condition in which the proton gradient rapidly dissipates, while the ATP/ADP+Pi ratio remains high. Under these conditions, FoF1 initially catalyzes ATPase-dependent proton pumping back to the lumen, the reverse of synthase activity. This wasteful consumption of the energy captured in light is rapidly halted by formation of a disulfide bond in the g subunit that stops rotation and ATP hydrolysis (6). Upon illumination, the first reducing equivalents from PSI that reduce ferredoxin are diverted from FNR to thioredoxin reductase that catalyzes the reduction of thioredoxin. Thioredoxin in turn reduces the disulfide of the g subunit to activate FoF1.